Thursday, July 06, 2006
Thoughts on the "Lost Common Ancestor"
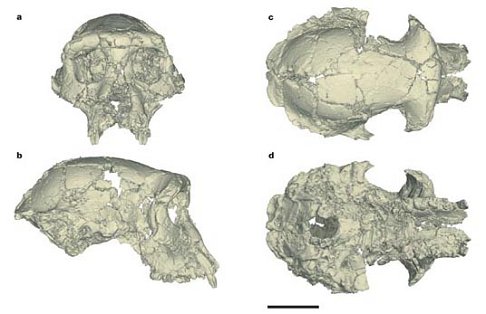
Just saw the Science Channel documentary, “Pre-human: Riddle of the Skull.” It’s the first attempt to popularize the discovery and explain the significance of the Sahelanthropus cranium. For those of you who need a reminder, a remarkably well-preserved, but distorted cranium of a hominoid dubbed Sahelanthropus tchadensis was discovered in 2001 in the North African country of Chad by a French paleontological team led by M. Brunet. The video reviewed the evidence, based on faunal studies, that Toumai, as the specimen has been nicknamed, is between 6 and 7 million years old. In the video Brunet, and various colleagues, argue that Toumai, is a full-fledged human ancestor, a hominid in the classic sense of the term, and as such the oldest and most primitive member of the “human family.” There are, however, some real problems with this assessment.
First, let’s look at some of the evidence they put forth regarding the “hominid” (or hominin if you prefer) status of Toumai. This includes aspects of cranial anatomy elucidated by a virtual reconstruction conducted by a team including D. E. Lieberman and D. Pilbeam, who were prominently interviewed in the video. The reconstruction demonstrates that the nuchal plane of the occiput, i.e. the surface of the skull base where the neck muscles attach, is more horizontally oriented than in the great apes and within the range of variation of austrlopithecines such as “Lucy,” suggesting that Toumai was well-adapted to bipedalism. This combined with a relatively vertical face and reduced canines with apical wear (i.e. wear at the tip of the canine crown) are said to clearly place Sahelanthropus within the hominid camp. In contrast to these human-like features the mostly complete cranium has a very small brain (between 320 and 380 cc) comparable in size to that of chimpanzees and on the lower end of the australopithecine range. It also has massive brow ridges, suggesting that it may be a male, as well as two rooted pre-molars, likewise a primitive ape-like trait. All things considered. Brunet, et al. forcefully argue in the video that Toumai was a habitual biped, belonging to a species that was beginning to adapt to a new hominid lifestyle.
Actually a date of ~ 6 million years or younger for the LCA has been reported on repeatedly, based on multiple genetic analyses going all the way back to Vince Sarich’s pioneering immunological work in the late 60s. Sarich’s late divergence hypothesis sounded the death knell for Ramapithecus as an exclusive human ancestor precisely because the youngest “ramapithecine” fossils were too old and would have lived prior to the LCA. Sarich’s initial divergence date was calculated around 4 mya. More recently an overwhelming body of genetic evidence places the divergence date at no more than 6 million years. The sole holdouts for an early divergence date for the human/chimp split are Arnason et al., who posit a date of ~ 10.5 - 13 mya., based on MtDNA datasets that do not take into account mutation rate increases in primates and utilize non-primate calibration points. The implications of an early divergence for the chimp/human LCA are stated by Arnason et al. as follows:
The revised estimates of primate divergence times suggest a new hypothesis for primate evolution and dispersal: that the divergence between strepsirhines (lorises, lemurs) and anthropoids was contemporary with the break-up of Southern continents about 90 MYBP, with strepsirhines becoming isolated on Madagascar and later dispersing to Africa (and Asia) and anthropoids evolving in South America and subsequently colonizing Africa (and Asia), or possibly North America.
Those who accept an early date for the chimp/human divergence based on genetic evidence should accordingly adopt the above scenario as well. I doubt, however, that proponents of an early date for the human/chimp divergence will actively promote hypotheses of primate evolution in accord with what Arnason et al. propose, as they are totally inconsistent with mainstream thinking. Accepting early divergence dates for the chimp/human LCA thus amounts to cherry-picking a convenient date to support an interpretation of Sahelanthropus and other similar fossils as bona fide hominids (sensu stricto).
What then are the implications of accepting the genetic evidence for a late divergence of the chimp and human lineages? Well, to paraphrase Vince Sarich when he debunked the hominid status of Ramapithecus, “I don’t care what it looks like, if it’s more than 4 million years old it’s not a hominid.” Following the same logic, accepting the modern genetic evidence for a 6 million year old divergence date for the chimp/human LCA implies that Sahelanthropus, Orrorin, and early forms of Ardipithecus are not in the direct line of human ancestry. What then do they represent? Given the time range within which these fossils are found, the obvious conclusion is that they are representative of the LCA or a species very close to it! For all intents and purposes I propose that African hominoids from ~ 5.5 – 7.5 mya that show a mix of African ape and human features should be considered part and parcel of the population from which the human/chimp LCA was derived. To reiterate, Sahelanthropus, Orrorin and Ardipithecus kadabba should be considered to be the long sought after last common ancestor of the African Apes and humans. The hominid-like features of these fossils must, therefore, be the common inheritance of all African hominoids, including the chimpanzee and gorilla, as well as ourselves. This, of course necessitates that not only have humans evolved away from the LCA, but so have chimps and gorillas. But that should come as no surprise, since that is what evolution is all about. Just as the LCA was not a human, neither was it a chimp nor a gorilla. In other words, the African ape/human LCA apparently possessed many more human-like features in its locomotor adaptation, dentition and masticatory apparatus than most researchers currently accept.
One may legitimately ask, from whence did this human-like LCA come? The answer is also revelatory, as the logical candidate is not African but European. The best candidate for immediate ancestor of the African ape/human LCA is none other than the late Miocene Grecian hominoid Ouranopithecus macedoniensis, which has long been promoted as a true hominid (sensu stricto) by L. De Bonis and G. Koufos. Others have noted strong similarities between Ouranopithecus and both gorillas and chimps. Ouranopithecus is noted for its thick molar enamel, high degree of sexual dimorphism, with females having particularly human-like canines, and large gorilla like faces and extremely prominent brow ridges. The one nearly complete face of Ouranopithecus is thought to be a male, but it has relatively small canines no larger than a modern female gorilla's. It has a very well developed brow ridge, but again, it is not any larger than that of a modern female gorilla's, suggesting that modern African apes are hypertrophied relative to their Miocene ancestors.

Ouranopithecus is dated between 8 – 10 mya in Greece, at a time when the environment was very similar to what is later seen in the area of North Africa where Sahelanthropus was discovered. The extinction of hominoids in Europe during the late Miocene may have been coincident with their dispersal into North Africa as climatic zones shifted south.
Ouranopithecus, and its descendent Sahelanthropus, can be thought of as adapted to a mosaic environment of woodlands interspersed with grasslands, requiring a locomotor pattern emphasizing incipient bipedalism and heavy chewing associated with sclerophyllic (drought resistant) vegetation. In this way the African ape/human LCA would have had a suite of preadaptations (now referred to as exaptations) that became enhanced amongst the later australopithecines as climatic deterioration continued apace during the terminal Miocene/early Pliocene transition in north and northeast Africa. The ancestors of gorillas and chimpanzees continued to disperse further south, encountering more tropical environments, leading to a loss of the hominid-like features inherited from their late Miocene European and North African ancestors.
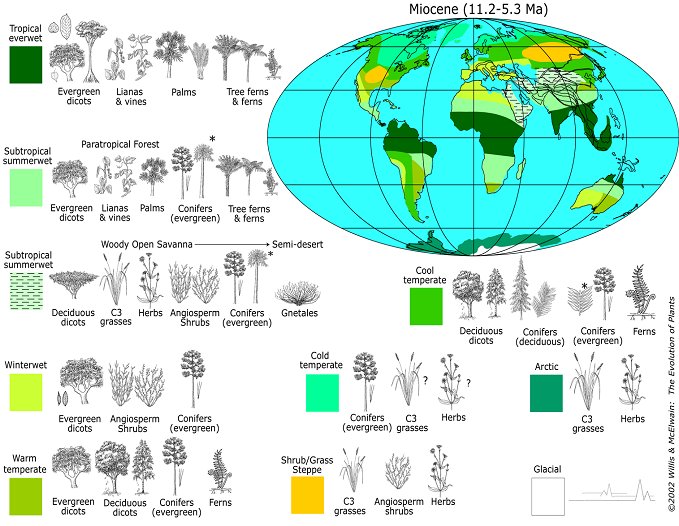 Late Miocene Vegetation Zones
Late Miocene Vegetation ZonesAccepting these ideas will require a major paradigm shift away from viewing human evolution as a result of a tropically adapted African ape acquiring hominid features as it expanded into or became isolated within increasingly harsh environments associated with late Miocene/early Pliocene climatic change. Human evolution is better seen as a continuation of trends established within European hominoid populations during the Miocene, which then were transferred south into northern Africa. North Africa then becomes a staging ground for the expansion of some populations of LCAs eastward into Ethiopia and then down the East Africa Rift Valley into South Africa, where they evolved into australopithecines and later human ancestors, and others populations which dispersed directly southward into equatorial Africa, producing the modern gorilla and chimpanzee.
Another consideration, brought to light by Reich et al., is that as populations of the LCA began to disperse, there would have been a considerable period of time when their descendents would not have diverged to the extent that they were genetically isolated from one another and hybridization could have potentially occured. This is illustrated in the chart below.
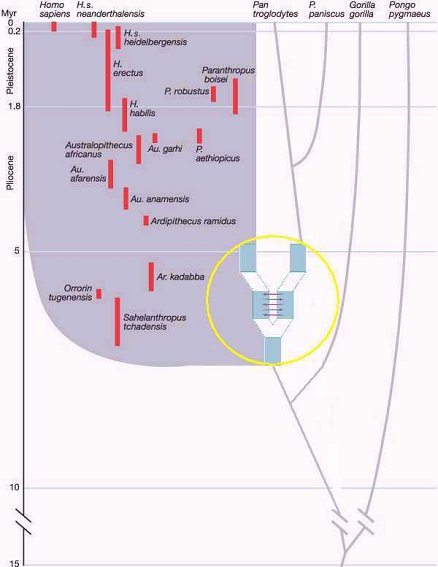
The fact is that early members of the human and chimpanzees lineages would have been nearly identical to one another for a considerable span of time. Their point of divergence can only really be discerned in hindsight. If Sahelanthropus is considered to be a hominid (hominin) it would imply that those adaptations that distinguish it from the coeval chimpanzee ancestor evolved in an extremely rapid fashion. The video thus portrays Toumai as very much a "hopeful monster" and human evolution as a dramatic departure from what had preceded it. I think it is much more realistic to view the first members of the human clade to be nearly indistinguishable from the LCA. In fact, it can be persuasively argued that the australopithecines, in preserving and enhancing many of the dietary and perhaps locomotor adaptations of late Miocene apes such as Ouranopithecus, were much more like the LCA than either the extant chimp or gorilla. In this manner, human evolution remains within the mainstream of hominoid evolution and is not some aberrant evolutionarily contingent phenomenon.

<< Home
![]()
