Sunday, July 30, 2006
Check-out This Blog's Sister Site
I've just recently updated the sister site to this blog, "The Fossil Evidence for Human Evolution in China." (N.B. the prior url for the site - www.chineseprehistory.org - was hijacked by a commercial outfit much to my chagrin). I've added a link to "Sinathropus" and a video file of a Chinese newscast reporting on the reconstruction of the Yunxian 2 cranium of Homo erectus that my colleague Prof. Li Tianyuan and I described back in the early 1990s. The site has a catalogue of Chinese hominid discoveries and many other resources relating to Chinese paleoanthropology, including pdfs of articles I've written on the subject, so check-it out if you haven't already.
Saturday, July 29, 2006
Hopping Across Laurasia
My previous post introduced readers to the fascinating research of Smith et al. regarding the dispersal of the earliest known euprimate genus Teilhardina from its apparent birthplace in China across the northern hemisphere to North America. Smith et al's. research is based on a high resolution assessment of the sedimentary environments in which the fossils were found. New investigative techniques allowed them to determine whether the fossil-bearing deposits were laid down before, during or after a climatic maximum that occured near the Paleocene/Eocene transition ~ 55 mya. A sequence based on carbon isotope ratios showed that Teilhardina asiatica was earliest, followed by T. belgica and T. americana . The spread of Teilhardina out of Asia into first Europe and then North America, across what was the more or less contiguous supercontinent of Laurasia, happened at a time when global warming is hypothesized to have spread a continuous canopy of evergreen forest across the northern latitudes. Many other mammalian genera are documented to have followed a similar eastward dispersal route.
Studies of this sort are what makes paleontology such an exciting field and shows it to be an experimental science in which hypotheses are generated by preliminary analysis of data collected in the field and then tested by falsifiable experimentation in the lab. There is one "true" story of the evolution and spread of life on earth which we are constantly unraveling with new and ever more analytic techniques.
Below is a figure from Smith et al's PNAS article that illustrates the research they conducted:
 Paleogeographic map showing hypothetical migration routes of Teilhardina during the earliest Eocene. (A–C) Timing of migration is obtained by correlations of the
Paleogeographic map showing hypothetical migration routes of Teilhardina during the earliest Eocene. (A–C) Timing of migration is obtained by correlations of the  13C excursion in North America (A), Europe (B), and Asia (C). T. brandti and other modern mammals first occur in the Lower Double Red (1,512 m) of Polecat Bench, Wyoming, which is situated above the minimum value of the 13C excursion (spike at 1,507 m). The Lower Double (L.D-) Red has an estimated age of 19–25 Kyr above the P/E boundary, based on soil carbonate nodules (SNC) and dispersed (disp.) organic carbon (DOC) (14, 20, 21). U.D-, Upper Double; Purp., purple; Fm, formation. (D) We hypothesize that Teilhardina dispersed through continuous forest from southern Asia to Europe and from nothern Europe to North America during the first 25 Kyr of the PETM.
13C excursion in North America (A), Europe (B), and Asia (C). T. brandti and other modern mammals first occur in the Lower Double Red (1,512 m) of Polecat Bench, Wyoming, which is situated above the minimum value of the 13C excursion (spike at 1,507 m). The Lower Double (L.D-) Red has an estimated age of 19–25 Kyr above the P/E boundary, based on soil carbonate nodules (SNC) and dispersed (disp.) organic carbon (DOC) (14, 20, 21). U.D-, Upper Double; Purp., purple; Fm, formation. (D) We hypothesize that Teilhardina dispersed through continuous forest from southern Asia to Europe and from nothern Europe to North America during the first 25 Kyr of the PETM.
Friday, July 28, 2006
New Evidence for the Asian Origins of Primates
A recent study by University of Michigan paleontologists Thierry Smith, Ken Rose and Philip Gingerich in the Procedings of the National Academy of Sciences has given unprecedented temporal resolution to the biogeographic first appearence of early Eocene primates. The research centers on the primate genus Teilhardina, recognized as one of the first true primates to appear in the fossil record. It was first discovered in Belgium in 1927 and originally described as belonging to the fossil genus and species Omomys belgicus by famed theologian and paleontologist Teilhard de Chardin. In 1940 Gaylord Simpson attributed the material to a new genus and species given the honorific nomen Teilhardina belgica. In 1970 a similar species, Teilhardina americana was described from early Eocene deposits in Wyoming. Then in 2004 an Asian form, Teilhardina asiatica, was discovered from the earliest Eocene (~55 mya) of the Hengyang Basin, China.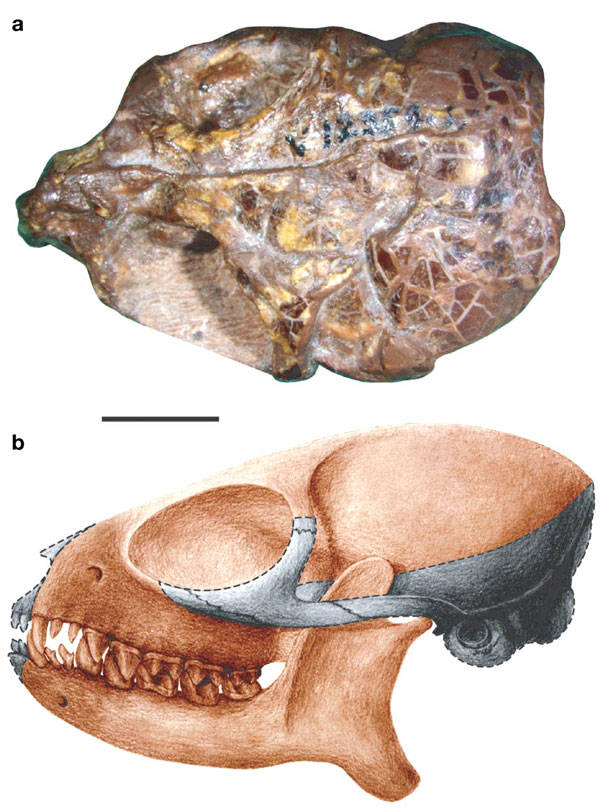
The skull of Teilhardina asiatica sp. nov. (IVPP V12357). a, Dorsal view of the skull. b, Reconstruction of the skull based on IVPP V12357, with grey shadow indicating the missing parts. Scale bar, 5 mm.
The earliest true primates (i.e. euprimates) have been sorted into two seperate groups; the omomyids and adapids. The omomyids have long been thought to be ancestral to later haplorhine primates (tarsiers and anthropoids), while adapids have been seen as broadly ancestral to later strepsirhine primates (the living lemurs and lorises). As the most primitive known omomyid, Teilhardina appears to be close to the last common ancestor of these two early groups of euprimates. Teilhardia seems to fit in near the base of the primate family tree close to numeral 2 on the figure above
Teilhardia seems to fit in near the base of the primate family tree close to numeral 2 on the figure aboveWhat Smith et al. have succeded in doing is sequencing the first appearence of the three regional species of Teilhardina. This has been determined by studying the carbon isotope ratios of deposits that have yielded the fossils in their respective regions. According to a UM press release:
To achieve such precision, they used a carbon isotope curve recently documented on all three continents. Carbon in the atmosphere, earth and living organisms differs in the proportion of carbon-12 and carbon-13 present. A flood of carbon- 12 is associated with the onset of an event known as the Paleocene-Eocene thermal maximum (PETM), one of the most rapid and extreme global warming events recorded in geologic history. It was during the PETM that modern primates first appeared 55 million years ago. Teilhardina in Asia precedes the maximum flood of carbon-12, Teilhardina in Europe coincides with it, and Teilhardina in North America appears just after the maximum.The most likely scenario has Teilhardina, and hence the omomyids in general, originating in Asia and quickly dispersing first to Europe and then across northern Laurasia to North America. The earliest adapid primate, Cantius, is currently known only in Europe and North America from slightly younger deposits than Teilhardina. The presense of adapids in Asia is problematical and still largely unresolved. Nevertheless, the current study confirms that the earliest true primate in the fossil record is from China, pointing to a possible Asian origin for the euprimate radiation.
Is there other evidence pointing to an Asian origin for primates? In a word, yes. Recent molecular studies indicate that modern placental mammals can be divided into four major clades: Afrotheria, Laurasiatheria, Xenarthra and Euarchontaglires. These groups seem to have biogeographic significance. Afrotheres include living orders of mammal that are primarily indigenous to Africa, such as elephants, manatees, hyraxes, aardvarks and some small insectivorous forms. Xenarthrans are the sloths, giant anteaters and armadillos indigenous to South America. Laurasiatheres include virtually all other placental mammals (various ungulates, cetaceans, bats, carnivorans, insectivores, etc.) and seem to have had an origin in the northern hemisphere Laurasian supercontinent spanning North America Europe and Asia. Euarchontaglires is the most interesting of these supraordinal mammalian clades from an anthropocentric perspective. It includes primates as well as the closely related tree shrews (Scandentia) and flying lemurs (Dermoptera) in the subgroup Archonta, and rodents and rabbits in the subgroup Glires. The extant archontans, other than primates, i.e. the tree shrews and flying lemur are found exclusively in continental and insular southeast Asia. Rodents and Lagomorphs (rabbits, hares and pikas) are global in distribution, but some of the earliest rodents are known from the Chinese Paleocene, as are the anagalid ancestors of lagomorphs. The biogeography of extant and extinct members of the euarchantaglires clade thus suggests East Asian and/or southeast Asian origins. Another indicator of East Asian origins for the primates, in particular, is the presense of the earliest anthropoid-like primates, the eosimiids, from the middle Eocene of China.
As things now stand a persuasive argument, based on biogeographic, molecular and fossil evidence, can be made for the Asian origins of euprimates, as well as other euarchontaglires, during the late Paleocene.
Sunday, July 23, 2006
Early Pleistocene Asian Hominid Redux
The recent surge of interest in possible Asian origins for the genus Homo prompts me to republish a piece I wrote a decade ago regarding what was then known of the earliest Asian hominins. Back then it was widely accepted by proponents of the “Out of Africa” hypothesis that Homo erectus was an evolutionary side branch of the human family tree, restricted to Asia, that eventually went extinct without issue.
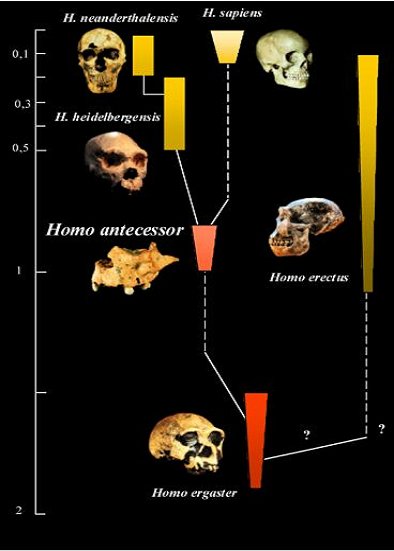 Popular "Out of Africa" phylogeny that isolates Homo erectus as a uniquely Asian species
Popular "Out of Africa" phylogeny that isolates Homo erectus as a uniquely Asian speciesOur lineage was universally accepted to have evolved in Africa from descendents of H. ergaster, which was considered to be the last common ancestor of all later hominins, including both H. erectus and H. sapiens. In this scenario H. ergaster diverged into an Afroeuropean lineage, which subsequently split into European Neanderthals and the African ancestors of modern H. sapiens, and an Asian lineage, comprising H .erectus and its localized descendents in East and Southeast Asia. The hunt was thus on for early antecedents of an exclusively Asian H.erectus.
With the discovery of relatively complete hominin cranial remains circa one million years BP in Ethiopia and Europe that are nearly indistinguishable from penecontemporaneous classic H. erectus from China and Java, the idea of an exclusively Asian H. erectus distinct from an ancestral East African H. ergaster has, however, begun to recede. The acceptance of H. ergaster as a distinct hominin taxon is no longer as widely accepted as previously and H. erectus is now generally recognized as the pandemic early Pleistocene ancestor of all later hominins, including modern humans.
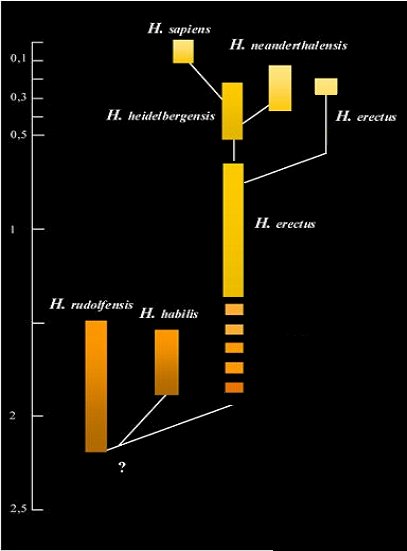
Widely accepted hominin phylogeny in which H. erectus is seen as the common ancestor of all later humans with a late survival in Asia as a relic species
As discussed in previous posts the discovery of the Dmanisi hominins dated to ~1.7 mya in the Republic of Georgia, has prompted some to suggest that the genus Homo had an extra-African origin. Is there really any substantial evidence for this hypothesis besides that presented and discounted in my previous posts on the Dmanisi remains? Not much new has emerged in the fossil record of China or Java relating to the presence of Plio-Pleistocene hominins (aka hominids) since I wrote the article mentioned at the top of this post although D. Tyler et al have recently described fragmentary and heavily distorted fossil remains attributed to “Meganthropus.” I will comment of that material in due course. Meanwhile, please proceed to my article on Plio-Pleistocene Asiatic hominids presented here.
Friday, July 21, 2006
Lessons from Supposed Anglo-Saxon Apartheid
An interesting study by Dr Mark Thomas et al., of the University College, London, Department of Biology et alia, suggests that, “The native Britons were genetically and culturally absorbed by the Anglo-Saxons over a period of as little as a few hundred years." A UCL press release on the research can be found here.
The study, which appears in the recent edition of the Proceedings of the Royal Society B: Biological Sciences, attempts to reconcile historic and archeological evidence for a limited incursion of Anglo-Saxons during their invasion and occupation of Britain, with genetic evidence showing the wholesale replacement of indigenous Celtic Y-chromosomes with those derived from a continental German substrate. As the article's abstract states:
The role of migration in the Anglo-Saxon transition in England remains controversial. Archaeological and historical evidence is inconclusive, but current estimates of the contribution of migrants to the English population range from less than 10000 to as many as 200000. In contrast, recent studies based on Y-chromosome variation posit a considerably higher contribution to the modern English gene pool (50–100%). Historical evidence suggests that following the Anglo-Saxon transition, people of indigenous ethnicity were at an economic and legal disadvantage compared to those having Anglo-Saxon ethnicity. It is likely that such a disadvantage would lead to differential reproductive success. We examine the effect of differential reproductive success, coupled with limited intermarriage between distinct ethnic groups, on the spread of genetic variants. Computer simulations indicate that a social structure limiting intermarriage between indigenous Britons and an initially small Anglo-Saxon immigrant population provide a plausible explanation of the high degree of Continental male-line ancestry in England.While interesting in and of itself, the study highlights that rapid genetic replacement can occur when two populations come into sustained contact, with hard to document demographic factors leading to fundamental shifts in the genetic composition of descendent populations.
This has a direct bearing on the controversy surrounding the role of Neanderthals in the descent of modern Europeans. My previous post hails the announcement that a concerted attempt has been initiated to completely decode the Neanderthal genome. This will eventually allow scientists to determine with certainty the contribution, if any, that Neanderthals made to modern European populations.
No matter what the ultimate outcome of these future studies may be, the issue of genetic replacement must take into account demographic factors such as those discussed in the research described above. There is no question that modern humans replaced the more archaic Neanderthal population of Europe in a relatively short span of time (~ 8000 year). The dynamics of this replacement are still debated and should be the focus of future research (see here, abstract A28 for example). Less emphasis should be placed on subjective questions of whether or not Neanderthals were a separate species of humankind.
Imminent Sequencing of the Neanderthal Genome
In approximately two years time what will be perhaps the seminal event of the 21st century will take place. The complete Neanderthal nuclear genome will be sequenced in a collaborative effort by the Max Planck Institute for Evolutionary Anthropoloy in Leipzig, Germany, and 454 Life Sciences Corporation, in Branford, Connecticut. Prof Svante Pääbo, Director of the Institute's Department of Evolutionary Anthropology, and Dr. Michael Egholm, Vice-President of Molecular Biology for 454 Life Sciences will jointly direct the project, made possible by financing from the Max Planck Society. 454 Life Sciences' newly developed sequencing technology has made it possible to extract and sequence nuclear DNA from fossil bone, a hopeless task using traditional techniques.
The significance of sequencing the nuclear genome of Neanderthals cannot be overestimated. Not only will it definitively demonstrate the nature of their relationship to modern humans, it will also define what it is that makes us distinct from our immediate pre-modern ancestors. Combined with the sequencing of the Chimpanzee genome we will be in the position to understand, to an unprecedented degree, the genetic transformation from ape to human. The implications for medical research are also stupendous, as we will be able to hone in on significant genetic sequences unique to modern people and compare them to both pre-modern human and pre-human sequences. As the techniques employed become more sophisticated the prospects for unraveling other aspects of our evolutionary past will increase exponentially. We will eventually be able to infer the nuclear genome of the chimpanzee/human last common ancestor and the genomes of ancestors throughout the course of human evolution. What an exciting prospect! Read more details about the project and view a webcast press conference about it from this Max Planck Institute press release and learn more about the techniques involved from this press kit by 454 Life Sciences.
Wednesday, July 12, 2006
Problems with “Out of Savannahstan”
A recent story in the popular science magazine “New Scientist” by Marek Kohn (Made in Savannahstan, NS July 1-7, 2006, pp 34-39) reviews a provocative article by Robin Dennell of the University of Sheffield, UK, and Wil Roebroeks of Leiden University, the Netherlands that appeared in the British journal “Nature” last December.
Dennell and Roebroeks hypothesize that hominins from the remakable Late Pliocene (~1.7 mya) Dmanisi site in the Republic of Georgia are the descendents of a yet unknown group of proto-humans that had previously dispersed into a belt of savannah extending from northern Africa across into southern Asia. It is this stock that adapted to the rigors of a grasslands environment and gave rise to Homo erectus, which then, much like the prodigal son, returned to Africa to smite its primitive cousins, the australopithecines (including early members of the genus Homo such as H. habilis and H. rudolfensis which they believe were advanced australopitecines.)
A linchpin of this scenario is the placement of australopithecines in Chad, far to the west of their previously known range of distribution, 3 to 3.5 mya, suggesting that australopithecines were capable of wide-ranging dispersals. This, in combination with early dates (> 1.5 mya) for Indonesian H. erectus from Sangiran and Mojokerto led the authors to speculate that there was an early dispersal of australopithecines out of Africa into Asia, where they were subjected to intensive selection resulting in the evolution of H. erectus. The sudden appearance of H. erectus in East Africa commencing ~ 1.8 mya, replacing species of so-called “early Homo,” was the result of their subsequent redeployment back into Africa.
There are a number of problems with the underlying premises of this “Out of Asia” hypothesis for the origins of Homo erectus. First, the presence of australopithecines in Chad is not unexpected given the recent discovery of 7 million year old Sahelanthropus in the same country. While I do not accept the hominin (aka hominid) status of Sahelanthropus, I do recognize it as a likely candidate for “Last Common Ancestor” of the African apes and humans (see previous post). As I see it, descendents of Sahelanthropus that stayed in northern Africa and dispersed east into Ethiopia and throughout the Rift Valley evolved into later australopithecines, while separate descendent lineages of Sahelanthropus that dispersed southward into equatorial Africa evolved into Gorillas and Chimps. In this scenario the presence of australopithecines in north central Africa at the time in question (3-3.5 mya) is to be expected, as it’s part of their ancestral homeland.
The small brain size, archaic facial morphology and Oldowan like tools of the Dmanisi hominins are also no surprise if one is familiar with fossil hominins from East Asia. I have previously favorably compared the Lantian skullcap from Gongwangling in China (dated at ~ 1.2 mya) with early H. erectus from East Africa. The Gongwangling calotte is estimated to have had a small cranial capacity (~ 780 cc) and a less derived facial morphology than later Chinese or Indonesian H. erectus. The hypothesis that there was a dispersal of an early grade of H. erectus out of Africa commencing ~ 1.8 mya is thus consistent with all available evidence and does not, at this time, require any premature modification. The quest for australopithecine fossils in Asia has been Quixotic at best and until there is some physical evidence for their presence outside of Africa hypotheses such as those of Dennel and Roebroeks will find only limited and I would suggest, ill-informed support (the presence of a "habiline" hominin at the Longgupo site in Sichuan, China dated to ~ 1.9 - 2.0 mya as proof of an early dispersal of pre-erectus hominins into Asia has been critiqued and is no longer generally accepted).
The article by Kohn makes reference to some preconceptions concerning the dispersal of H. erectus out of Africa perpetuated by commentators such as Ian Tattersall. One such idea is that humans expanded their range into Asia, or vice versa, due to a “typically insatiable human wanderlust.” This is just plain silly. Species, whether human or not, do not consciously decide to strike out on transcontinental treks, they expand into adjacent territory that they are equipped to exploit, both physically and behaviorally. Humans, incidentally, were not the first hominoids to disperse throughout the Old World. Did late Miocene apes, which ranged from Spain to China, have an “insatiable wanderlust?” No, they had the behavioral and physical adaptations that enabled them to exploit contiguous environments spread across the southern tier of Eurasia. This is, obviously, how all-major dispersals take place and humans are no different from other species in this regard.
If, as I have suggested in previous posts, the australopithecines are the lineal descendents of late Miocene European apes that were displaced southwards into northern Africa at the end of the Miocene because of climatic and environmental deterioration in their native clines, it would appear that the southern tier of Eurasia was inhospitable to large-bodied hominoids for a considerable span of time. Northern and northeastern Africa, which retained the open woodlands environment that suited these hominoids, was the cauldron in which the australopithecines evolved. As this environment slowly dried out and was replaced by expanding grasslands, robust australopithecines and the ancestors of the genus Homo diverged. The adaptations forged by early Homo for life on the savannah allowed for range expansion as soon as they emerged, hence the rapid dispersal of early Homo, commencing around 1.9 mya into the Middle East and the Caucus region and rapid subsequent eastward expansions into east and southeast Asia. This scenario is in keeping with the broad outline of human evolution generally accepted by most contemporary anthropologists and does not need any major revision based on presently available evidence.
As Templeton has documented using genetic data, there were apparently multiple dispersals out of Africa beginning ~ 1.9 mya. This was coupled with a considerable amount of back migration and multidirectional genetic admixing throughout the latter phases of human evolution. What the Dmanisi remains demonstrate is that between 1.7 and 1.9 mya there was what appears to have been a rapid transition from a “habiline to an “erectine” grade of hominin evolution. It is not surprising to find specimens during this period that show different mixes of primitive and derived features. What we’re witnessing may be the human lineage in the throes of evolutionary change as new genetic combinations are sorted out and eventually become channelized into the morphotype of classic Homo erectus, a pattern that once established remained virtually unchanged for nearly one million years.
Part of this transition was the development of new stone tool fabrication techniques, eventually leading to the emergence of a full-fledged Acheulean tradition by ~ 1.65 mya in East Afria. Earlier Oldowan-like traditions now appear to have spread into Eurasia prior to the spread of more sophisticated hand-axe traditions. But the idea that early Homo needed Acheulean tools and large brain size to expand their range out of Africa was never a viable hypothesis. For decades it was noted that Chinese and Indonesian H. erectus lacked major components of the Acheulean tradition and the so-called Movius line was established to separate the early Paleolithic into eastern and western traditions. The stone tool industries of Asian H. erectus were referred to as part of a chopper/chopping tool complex of a basically Oldowan nature. Early H. erectus in Asia is also noted for relatively small cranial capacities (i.e. the previously mentioned Lantian skullcap.). It’s been known for decades that hand-axes and large brains were not prerequisites for human expansion into the Far East.
In sum, australopithecines were not adapted to the savannah environments that were spreading across the southern tier of Eurasia at the end of the Miocene. It was the spread of these grasslands that spelled doom for the Eurasian hominoid antecedents of the African australopithecines. Africa seems to have been the incubator for the australopithecine adaptation to the woodland/grassland ecotone, a stable adaptation that remained little changed for nearly 2 million years (~ 4.5 – 2.5 mya). This environment does not seem to have prevailed in more northerly latitudes. As grasslands began sreading in Africa at ~ 2.5 mya generalized australopithecines seem to have split into robust lineages and those evolving towards the genus Homo. As of today genetic, archeological and paleontological evidence all points to an initial dispersal of Homo out of Africa at ~ 1.9 mya. Once this range expansion was achieved there seems to have been episodic genetic exchange between widely dispersed demes of an emerging H. erectus, the longest lasting hominin species and direct ancestor of all later humans.
Sunday, July 09, 2006
Comments on the Flores hominid
Gary Richards, an old friend and colleague from my days at the Laboratory for Human Evolutionary Studies (now the Human Evolution Research Center) at the University of California, Berkeley has a recent article in the Journal of Evolutionary Biology (DOI link) challenging the validity of Homo floresiensis. He argues that changes in developmental pathways of an isolated population of modern humans could have produced the idiosyncratic features seen in the Flores remains. Both John Hawks and Carl Zimmer offer useful reviews of Gary’s contribution, which can be found here and here.
I’ve always been skeptical of the view that the Flores hominid was an insular dwarf derived from an antecedent population of Indonesian H. erectus. The association of “H. floresiensis” with a modern human tool-kit and the presence of short-statured peoples throughout the region led me to suspect that if insular dwarfing was the cause for the features seen in the Flores remains, the process was acting on a population of modern humans rather than a relic population of H. erectus.
Gary’s article clearly shows that populational dwarfism among extant humans varies as to etiology and that the stature of LB1 does not fall too far below the lower limit of the range of variation seen in modern humans. Many of the seemingly “primitive traits” seen in the Flores remains may be due to allometric scaling effects or the result of alterations in developmental pathways brought on by changes in the expression of human growth hormones and associated factors. In addition, mutational effects leading to extreme size reduction, combined with inbreeding may have led to an increase in the incidence of microcephaly within individual demes of Flores hominids. This would help account for the unlikely recovery of an individual with such an extremely small cranial capacity as the LB1 specimen.
The article argues that more wide-ranging comparisons must be made between the Flores sample and localized human populations to better understand how “Homo floresiensis” fits into the panoply of human evolution and endemic adaptation. Gary concludes that:
When one applies the concept of maximum parsimony (Sober, 1988) to the totality of evidence available on the Flores remains, one finds significant support for the remains being a variant of H. sapiens and little support for a species-level distinction. Given this position, I suggest that the LB1–LB9 individuals represent the remains of a H. sapiens group which became dwarfed in an island environment via changes in the GH–IGF-I axis (Human Growth Hormone-Insulin-like Growth Factor I Axis). Acquisition of a dwarfing condition may either have occurred prior to or after the group arrived on the island. If it can be demonstrated that the totality of the recovered remains sample the same population, it appears that a mutation in the MCPH gene family or a secondary modification of the GH–IGF-I axis arose in the later part of their occupation of the island and was transmitted within a local group. Whereas I consider the ‘primitive’ features identified in the LB1–LB9 individuals to be consistent with the scenario presented above, only a detailed analysis will be able to clarify the value of these features for phylogeny reconstruction.
This seems to me to be an extremely well balanced analysis and one that addresses the major contradictions generated by the Flores remains.
Folkloric accounts by local residents of Floresian “hobbits” suggests that there was ongoing contact between the dwarfed population and settled villagers, leading to interbreeding and resultant genetic exchange between the two populations, again mitigating against “Homo floresiensis” being a distinct human species. No matter how one cuts it, however, the recovery of the Liang Bua remains expands the range of modern human variation to the brink of an incipient speciation event and helps document an extremely interesting episode in recent human evolution.
Saturday, July 08, 2006
More on the "Lost Common Ancestor"
Happened upon John Hawks website cum blog, which has a recent post regarding Sahelanthropus. Hawks has had a long standing collaboration with Milford Wolpoff, and he, Wolpoff, Brigitte Senut, Martin Pickford and James Ahern have published a critique of Toumai as an ancestral hominid. Interestingly enough, Senut and Pickford have made claims about the hominid status of Orrorin tugenensis, a 6 million year old hominoid from the Turgen Hills of Kenya, similar to those made by Brunet et al. about Sahelanthropus. Orrorin, discovered in 2000 and nicknamed "millenium man," consists of a proximal femur and other fragmentary skeletal and dentognathic remains, that have been described as "the first human" by Pickford and Senut. As the search for the "first human" is a very competitive business, it is not surprising that Pickford and Senut concur with Wolpoff et al's analysis of Sahelanthropus as an ape, although they differ in their interpretation as to its overall affinities. Pickford and Senut's insistance that Orrorin was a bipedal hominid with a modern human gait, has also been challenged, so there is a certain irony in their collaboration with Wolpoff et al. in deconstructing Sahelanthropus.
Although there is a general willingness to accept Orrorin as a biped and hence an early hominid, the nature of its bipedalism has been called into question, with White et al. suggesting that the anatomy of the femur was more primitive than seen in australopithecines. Of course bipedalism in late Miocene hominoids has been argued for some unequivocally non-hominid apes such as the 8 million year old remains of Oreopoithecus from Tuscan lignites in Italy, so bipedalism in and of itself should not necessarily serve as the sole criterion for being human. As I argued in my last post, faculative bipedalism may have been part of the morphotype of the African ape/human LCA and not indicative of true homind status. It is interesting that no one seems to be arguing for this interpretation. It's either argued that a six million year old divergence date for the human/chimp split is too recent or that the fossils in question do not possess the features they are said to possess and are not really hominids after all. This is exemplified by a quote of Vince Sarich's from an article in Science by Ann Gibbons:
The first molecular study back in 1967 dated the split between
humans and apes to 5 million years ago, and Vince Sarich of the University
of California, Berkeley, co-author of the study, still stands by
that date. “I still bet that either the morphology or dates or both will
be found wanting for these ‘6-million-year-old hominids,’ ” he says.
The idea that the LCA of African apes and humans may have had preadatations for bipedalism that are expressed morphologically as early signs of a bipedal adaptation, does not seem to have been entertained by anyone besides yours truely. I agree with White et als. contention that Sahelanthropus, Orrorin and Ardipithecus represent the same taxon, but differ as to their interpretation that it is a hominid. As I said in my last post all three should be recognized as representative of the "lost" common ancestor of chimps and humans, a view consistent with their chronometric age and the consensus 5-6 million year old divergence date for the LCA established by genomic analysis. Again let me reiterate, because it seems to be a point no one is willing to concede, based on current evidence, the most parsimonious position regarding the phylogenetics of Sahelanthropus, Orrorin, and early forms of Ardipithecus is that they are indistinguishable from the human/chimp LCA and that this LCA possessed more hominid-like features vis a vis its locomotor and dietary adaptations than anyone has been willing to acknowledge.
Thursday, July 06, 2006
Thoughts on the "Lost Common Ancestor"
Just saw the Science Channel documentary, “Pre-human: Riddle of the Skull.” It’s the first attempt to popularize the discovery and explain the significance of the Sahelanthropus cranium. For those of you who need a reminder, a remarkably well-preserved, but distorted cranium of a hominoid dubbed Sahelanthropus tchadensis was discovered in 2001 in the North African country of Chad by a French paleontological team led by M. Brunet. The video reviewed the evidence, based on faunal studies, that Toumai, as the specimen has been nicknamed, is between 6 and 7 million years old. In the video Brunet, and various colleagues, argue that Toumai, is a full-fledged human ancestor, a hominid in the classic sense of the term, and as such the oldest and most primitive member of the “human family.” There are, however, some real problems with this assessment.
First, let’s look at some of the evidence they put forth regarding the “hominid” (or hominin if you prefer) status of Toumai. This includes aspects of cranial anatomy elucidated by a virtual reconstruction conducted by a team including D. E. Lieberman and D. Pilbeam, who were prominently interviewed in the video. The reconstruction demonstrates that the nuchal plane of the occiput, i.e. the surface of the skull base where the neck muscles attach, is more horizontally oriented than in the great apes and within the range of variation of austrlopithecines such as “Lucy,” suggesting that Toumai was well-adapted to bipedalism. This combined with a relatively vertical face and reduced canines with apical wear (i.e. wear at the tip of the canine crown) are said to clearly place Sahelanthropus within the hominid camp. In contrast to these human-like features the mostly complete cranium has a very small brain (between 320 and 380 cc) comparable in size to that of chimpanzees and on the lower end of the australopithecine range. It also has massive brow ridges, suggesting that it may be a male, as well as two rooted pre-molars, likewise a primitive ape-like trait. All things considered. Brunet, et al. forcefully argue in the video that Toumai was a habitual biped, belonging to a species that was beginning to adapt to a new hominid lifestyle.
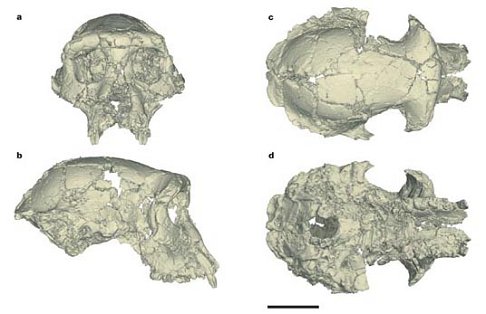
Actually a date of ~ 6 million years or younger for the LCA has been reported on repeatedly, based on multiple genetic analyses going all the way back to Vince Sarich’s pioneering immunological work in the late 60s. Sarich’s late divergence hypothesis sounded the death knell for Ramapithecus as an exclusive human ancestor precisely because the youngest “ramapithecine” fossils were too old and would have lived prior to the LCA. Sarich’s initial divergence date was calculated around 4 mya. More recently an overwhelming body of genetic evidence places the divergence date at no more than 6 million years. The sole holdouts for an early divergence date for the human/chimp split are Arnason et al., who posit a date of ~ 10.5 - 13 mya., based on MtDNA datasets that do not take into account mutation rate increases in primates and utilize non-primate calibration points. The implications of an early divergence for the chimp/human LCA are stated by Arnason et al. as follows:
The revised estimates of primate divergence times suggest a new hypothesis for primate evolution and dispersal: that the divergence between strepsirhines (lorises, lemurs) and anthropoids was contemporary with the break-up of Southern continents about 90 MYBP, with strepsirhines becoming isolated on Madagascar and later dispersing to Africa (and Asia) and anthropoids evolving in South America and subsequently colonizing Africa (and Asia), or possibly North America.
Those who accept an early date for the chimp/human divergence based on genetic evidence should accordingly adopt the above scenario as well. I doubt, however, that proponents of an early date for the human/chimp divergence will actively promote hypotheses of primate evolution in accord with what Arnason et al. propose, as they are totally inconsistent with mainstream thinking. Accepting early divergence dates for the chimp/human LCA thus amounts to cherry-picking a convenient date to support an interpretation of Sahelanthropus and other similar fossils as bona fide hominids (sensu stricto).
What then are the implications of accepting the genetic evidence for a late divergence of the chimp and human lineages? Well, to paraphrase Vince Sarich when he debunked the hominid status of Ramapithecus, “I don’t care what it looks like, if it’s more than 4 million years old it’s not a hominid.” Following the same logic, accepting the modern genetic evidence for a 6 million year old divergence date for the chimp/human LCA implies that Sahelanthropus, Orrorin, and early forms of Ardipithecus are not in the direct line of human ancestry. What then do they represent? Given the time range within which these fossils are found, the obvious conclusion is that they are representative of the LCA or a species very close to it! For all intents and purposes I propose that African hominoids from ~ 5.5 – 7.5 mya that show a mix of African ape and human features should be considered part and parcel of the population from which the human/chimp LCA was derived. To reiterate, Sahelanthropus, Orrorin and Ardipithecus kadabba should be considered to be the long sought after last common ancestor of the African Apes and humans. The hominid-like features of these fossils must, therefore, be the common inheritance of all African hominoids, including the chimpanzee and gorilla, as well as ourselves. This, of course necessitates that not only have humans evolved away from the LCA, but so have chimps and gorillas. But that should come as no surprise, since that is what evolution is all about. Just as the LCA was not a human, neither was it a chimp nor a gorilla. In other words, the African ape/human LCA apparently possessed many more human-like features in its locomotor adaptation, dentition and masticatory apparatus than most researchers currently accept.
One may legitimately ask, from whence did this human-like LCA come? The answer is also revelatory, as the logical candidate is not African but European. The best candidate for immediate ancestor of the African ape/human LCA is none other than the late Miocene Grecian hominoid Ouranopithecus macedoniensis, which has long been promoted as a true hominid (sensu stricto) by L. De Bonis and G. Koufos. Others have noted strong similarities between Ouranopithecus and both gorillas and chimps. Ouranopithecus is noted for its thick molar enamel, high degree of sexual dimorphism, with females having particularly human-like canines, and large gorilla like faces and extremely prominent brow ridges. The one nearly complete face of Ouranopithecus is thought to be a male, but it has relatively small canines no larger than a modern female gorilla's. It has a very well developed brow ridge, but again, it is not any larger than that of a modern female gorilla's, suggesting that modern African apes are hypertrophied relative to their Miocene ancestors.

Ouranopithecus is dated between 8 – 10 mya in Greece, at a time when the environment was very similar to what is later seen in the area of North Africa where Sahelanthropus was discovered. The extinction of hominoids in Europe during the late Miocene may have been coincident with their dispersal into North Africa as climatic zones shifted south.
Ouranopithecus, and its descendent Sahelanthropus, can be thought of as adapted to a mosaic environment of woodlands interspersed with grasslands, requiring a locomotor pattern emphasizing incipient bipedalism and heavy chewing associated with sclerophyllic (drought resistant) vegetation. In this way the African ape/human LCA would have had a suite of preadaptations (now referred to as exaptations) that became enhanced amongst the later australopithecines as climatic deterioration continued apace during the terminal Miocene/early Pliocene transition in north and northeast Africa. The ancestors of gorillas and chimpanzees continued to disperse further south, encountering more tropical environments, leading to a loss of the hominid-like features inherited from their late Miocene European and North African ancestors.
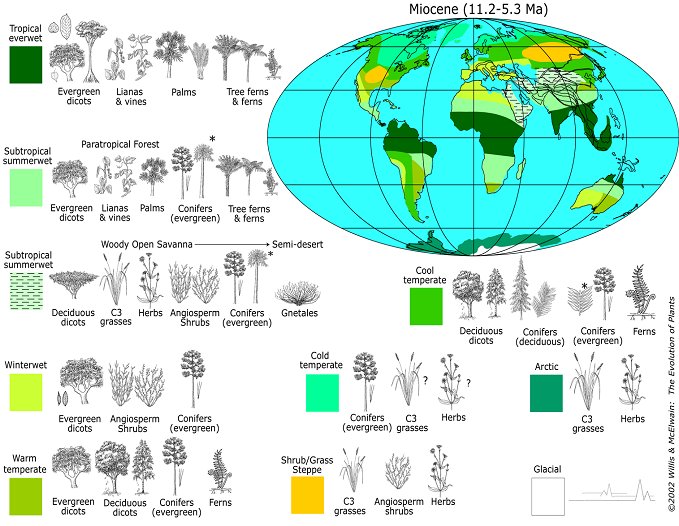 Late Miocene Vegetation Zones
Late Miocene Vegetation ZonesAccepting these ideas will require a major paradigm shift away from viewing human evolution as a result of a tropically adapted African ape acquiring hominid features as it expanded into or became isolated within increasingly harsh environments associated with late Miocene/early Pliocene climatic change. Human evolution is better seen as a continuation of trends established within European hominoid populations during the Miocene, which then were transferred south into northern Africa. North Africa then becomes a staging ground for the expansion of some populations of LCAs eastward into Ethiopia and then down the East Africa Rift Valley into South Africa, where they evolved into australopithecines and later human ancestors, and others populations which dispersed directly southward into equatorial Africa, producing the modern gorilla and chimpanzee.
Another consideration, brought to light by Reich et al., is that as populations of the LCA began to disperse, there would have been a considerable period of time when their descendents would not have diverged to the extent that they were genetically isolated from one another and hybridization could have potentially occured. This is illustrated in the chart below.
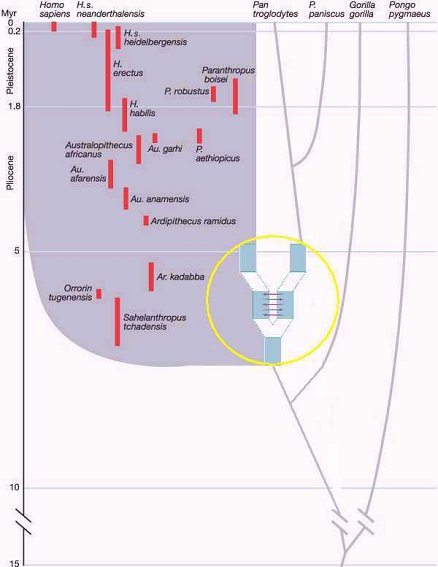
The fact is that early members of the human and chimpanzees lineages would have been nearly identical to one another for a considerable span of time. Their point of divergence can only really be discerned in hindsight. If Sahelanthropus is considered to be a hominid (hominin) it would imply that those adaptations that distinguish it from the coeval chimpanzee ancestor evolved in an extremely rapid fashion. The video thus portrays Toumai as very much a "hopeful monster" and human evolution as a dramatic departure from what had preceded it. I think it is much more realistic to view the first members of the human clade to be nearly indistinguishable from the LCA. In fact, it can be persuasively argued that the australopithecines, in preserving and enhancing many of the dietary and perhaps locomotor adaptations of late Miocene apes such as Ouranopithecus, were much more like the LCA than either the extant chimp or gorilla. In this manner, human evolution remains within the mainstream of hominoid evolution and is not some aberrant evolutionarily contingent phenomenon.
Saturday, July 01, 2006
Early Cambrian Echinoderms of China
As previous posts have shown the Lower Cambrian Chengjiang fauna of Yunnan, China has yielded a vast array of primordial animal fossils. I’ve focused on chordates and their allies as they are intrinsically interesting to us, their descendents. One of the more interesting non-chordates is an early echinoderm dubbed Vetulocystis catenat. PZ Myers over at Pharyngula has this post from 2004:
This week's Nature includes a wonderful description of an exotic group of deuterostomes from the lower Cambrian, 520 million years ago. Deuterostomes are animals characterized by their embryology: when they gastrulate, the site of closure of the migrating tissues, the blastopore, becomes the anus of the animal. This is in contrast to the protostomes, in which the blastopore becomes the mouth. We chordates are deuterostomes, as are echinoderms, some marine worms called hemichordates, and the urochordates, or sea squirts.
Shu et al. have identified some animals called vetulocystids, and they are most closely related to modern echinoderms. Echinoderm evolution is confusing and complicated, largely because the modern forms are so highly derived and distinct from the ancestral forms, and because the echinoderm lineage has been spectacularly diverse morphologically. The authors jump from some somewhat speculative interpretations of the fossil anatomy (the specimens, as you can see below, are peculiar and the structures difficult to identify) to an idea that clarifies the organization of the deuterostome lineage.
Other Early Chinese Fish-like Vertebrates and Pre-Vertebrates
As we go further back in time we encounter ever more ancestral organisms. In China precursors to the fish-like forms discussed in previous posts are found in the early Cambrian Chengjiang deposits from the southern province of Yunnan. A number of important early chordates and their allies have been described from these deposits. The vertebrates, which include mammals, birds, reptiles, amphibians and fish, have their origins within this assemblage. Clear fossils of primitive jawless fish (agnathans) have long been known from the Lower Ordovician Period (~475 million years ago), with more questionable examples from the earlier Cambrian Period (510-545 million years ago). Some of the newly described fossils from Chengjiang have been clearly identified as agnathans, a discovery that pushes back the fossil record of the vertebrates about 50 million years.
The specimens from Chengjiang include (descriptions from Vertebrate section of Palaeos and other sources):
Haikouella - originally thought to be a stem vertebrate, is now regarded as a Vetulicolian or other stem deuterostome. The Deuterostomes (meaning "secondary mouth") are a major animal taxon that includes the echinoderms (e.g., crinoids, starfish, sea urchins) and the chordates (e.g., sea squirts, lancelets, and vertebrates). The deuterostomes belong to a larger group within the Animalia called the Bilateria, owing to their bilateral symmetry along a median axis: note that some deuterostomes (i.e. echinoderms) are bilateral as embryos, later maturing to have radial symmetry. Shu & Conway Morris (2003) have made a point of "emphatically contest[ing] the vertebrate affinities of Haikouella, emphasizing the absence of eyes, notochord, myomeres, or brain.
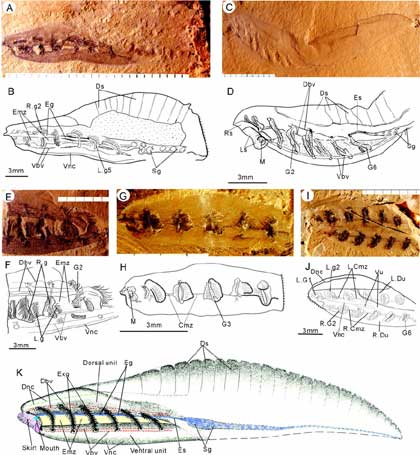
Examples of Haikouella and a reconstruction of H. jianshanensis. (A, B, and E to J) H. jianshanensis; (C and D) H. lanceolata. All are lateral views, except for (I) and (J). In (A)and (B), the dorsal posterior segments are faintly visible [specimen (226)]. The complete specimen with well-preserved dorsal segments (totaling 10) is shown in (C). A drawing of the anterior of (C) is shown in (D). The anterior with an expanded median zone [specimen (010)] is shown in (E) and (F), and the anterior with a closed median zone [specimen (358)] is shown in (G) and (H). An oblique view [specimen (238)] is given in (I) and (J). (K) A reconstruction of H. jianshanensis. Abbreviations are as follows: sal segments; Du, dorsal unit; Eg, ?epipharyngeal structure; Emz, expanded median zone; Es, esophagus; Exg, external gills; G1 to G6, gill arch 1 to 6; L.Cmz, left closed median zone; L.Du, left dorsal unit; L.g1 to L.g6, left gill arch 1 to left gill arch 6; Ls, left skirt; M, mouth; R.Cmz, right closed median zone; R.Du, right dorsal unit; R.g1 to R.g6, right gill arch 1 to right gill arch 6; Rs, right skirt; S, skirt; Sb, bar of skirt; Sbsb, space between skirt and body; Sg, ?spiral gut; Tmmz, thin membrane covering median zone; Vbv, ventral blood vessels; Vnc, ?ventral nerve cord; and Vu, ventral unit. Scale intervals are in millimeters.
Haikouella and related forms are classified as Yunnanozoans. They branched off the chordate or hemichordate lines very early on and are probably not far removed from the last common ancestor of deuterostomes as the diagram below suggests.

Haikouella must have been an important part of the Chengjiang biota as more than 1400 specimens were studied for the analysis of their characteristics as detailed in the Science article below.
Shu D, Conway Morris S, Zhang ZF, Liu JN, Han J, Chen L, Zhang XL, Yasui K, Li Y (2003) A New Species of Yunnanozoan with Implications for Deuterostome Evolution. Science 299(5611):1380-1384
Yunnanozoon lividum - from the Early Cambrian (about 530 million years ago) of Ma'anshan, Chengjiang, Yunnan, People's Republic of China has been described as the oldest known example of a hemichordate, but see above. The phylum (or 'supergroup') Hemichordata is closely associated with the phylum Chordata, which includes the vertebrates along with other groups such as cephalochordates and urochordates. As with Haikouella, the identification of Yunnanozoon is very significant for understanding the early evolution of chordates and their allies.

Yunnanozoon lividum
Haikouichthys - originally described by Shu et al. (1999) as one of two possible vertebrates on the basis of unique specimens from Chengjiang. The other taxon, Myllokunmingia, remains unique. It has been suggested that Myllokunmingia is the same as Haikouichthys, Hou et al. (2002). The issue of Myllokunmingia's exact characteristics and affinities became somewhat less urgent after five hundred new specimens of Haikouichthys were found near Haikou. Portions of the anatomy were then redescribed on the basis of this massive body of new data by an all-star cast of Chinese and Western early vertebrate specialists, Shu et al. (2002). They at first pronounced Haikouichthys to be not merely a chordate but a lamprey-like jawless fish, with gill bars supporting its gills. Myllokunmingia is said to have pouch-like structures associated with its gills, which also makes it a jawless fish. On the other hand a cladogram by Shu et al. (2003) identifies Haikouichthys as a basal chordate, or possibly a basal craniate, but not a vertebrate. Nevertheless, Haikouichthys has a number of characteristics thought to be derived features shared with true vertebrate. These discoveries emphasize the complexity of early vertebrate origins, but they do not change prevailing ideas about the evolution of fishes from earlier chordates. They do, however, change the timing of these transitions. It's now clear that the origin of vertebrate fishes is very early in the Cambrian. By the same token, the subsequent evolution of the fishes proceded at a much slower pace. It took about 30 m.y. before the cartilaginous structure of the earliest jawless fishes began to change into bone.
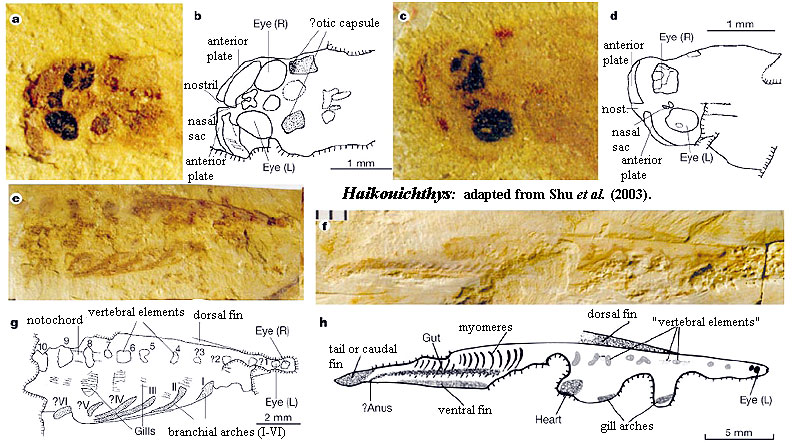
For more on early Chinese vertebrates such as Haikouichthys and Myllokunmingia see this article by R. Monastersky from Science News Online.
Cathaymyrus diadexus – also from the Early Cambrian at Ma'anshan, Chengjiang, Yunnan, People's Republic of China, is the oldest known example of a cephalochordate. The form of Cathaymyrus resembles Pikaia from the Middle Cambrian Burgess Shale of Canada, but this animal is about 10 million years older. Some paleontologists have suggested that the vertebrates, evolved from cephalochordates like Cathaymyrus.
 Cathaymyrus diadexus
Cathaymyrus diadexusXidazoon stephanus - from the Early Cambrian at Haikou, Kunming, Yunnan, People's Republic of China is more problematic as its relationship to the known animal groups is ambiguous. The anterior of this animal resembles an agnathan fish known as a pipiscid, but the segmented tail-like posterior is more like that of an invertebrate, such as an arthropod. Because of its enigmatic nature, Xidazoon has at present been assigned to the pseudo-phylum "Problematica". Nevertheless, this animal has great importance in the study of the evolution of the primitive deuterostomes and protostomes.

 Xidazoon stephanus
Xidazoon stephanus
Reconstruction is supervised by Prof. Shu Degan.
![]()

{kind=link}
{kind=link}
{kind=link}