Tuesday, June 22, 2010
Its time to sink the genus Australopithecus
With the recent description of the partial skeletons of Ardipithecus ramidus from Aramis and Australopithecus afarensis from Woranso-Mille, both in Ethiopia, a clear distinction can be made between pre-human and fully human adaptations to bipedality. When we include Au. anamensis in the equation, which is assumed by most to be a slightly older and more "primitive" version of Au. afarensis, the separation between Ardipithecus and Australopithecus is reduced to 200 ky, a mere blip on the paleoanthropological radar screen. The morphological distance is, however, far greater in both cranial capacity (less than 400 cc in Ardipithecus vs. more than 400 cc in Australopithecus) and overall locomotor pattern. To go from Ardipithecus to Australopithecus in such a short span of time seems hard to imagine (see previous post). On the other end of the temporal spectrum the recent description of Australopithecus sediba, dated to 1.9 mya from the Republic of South Africa, highlights the difficulty in separating late occurring australpiths from early members of the genus Homo. It thus seems, on the one hand, that adaptationally Australopithecus is much more similar to Homo than it is to Ardipithecus, while on the other hand, it is becoming more and more difficult to separate late occurring Australopithecus from early Homo. The rationale for maintaining the genus Australopithecus appears to be evaporating. It makes more sense to sink Australopithecus into the genus Homo while retaining it as a subgenus to distinguish it from later occurring, increasingly more human-like fossil hominins. For the sake of convenience, and other considerations, I think the genus Homo can be divided into four subgenera H. (Australopithecus), H. (Paranthropus), H. (Pithecanthropus), and H. (Homo). For example, “Lucy” would belong to H. (Australopithecus) afarensis, South African robusts to H. (Paranthropus) robustus, “Java Man” to H.(Pithecanthropus) erectus and “Heidelberg Man” to H. (Homo) heidelbergensis. Each one of these subgenera represents a clearly definable morphotype identifiable by a set of synapomorphies indicating a shared common ancestry as a "species clade" composed of distinct "paleo-demes" as discussed by Howell (1999). Colloquially we can refer to these constructs as australopiths, paranthropes, pithecanthropes, and archaics (i.e. pre-modern humans) vis-à-vis modern humans.
To quote the venerable resource Wikipedia, “in zoology, a subgeneric name can be used independently or included in a species name, in parentheses, placed between the generic and specific name … However, it is not mandatory, or even customary, when giving the name of a species, to include the subgeneric name.” Thus “Lucy” would commonly be referred to as H. afarensis, etc., etc. This is in partial conformity to the geneticist Morris Goodman’s suggestion that the African Great Apes be sunk into the genus Homo because of their overall genetic similarity to H. sapiens and J. T. Robinson’s suggestion that Au. africanus be revised to H. africanus. Goodman’s view, while biologically correct, seems to be a bit too radical for most people’s taste, while Robinson’s suggestion now appears to have been prescient. In the light of these precedents the sinking of Australopithecus into Homo, as a subgenus, seems eminently reasonable.
Monday, June 21, 2010
Ardipithecus fades into the past
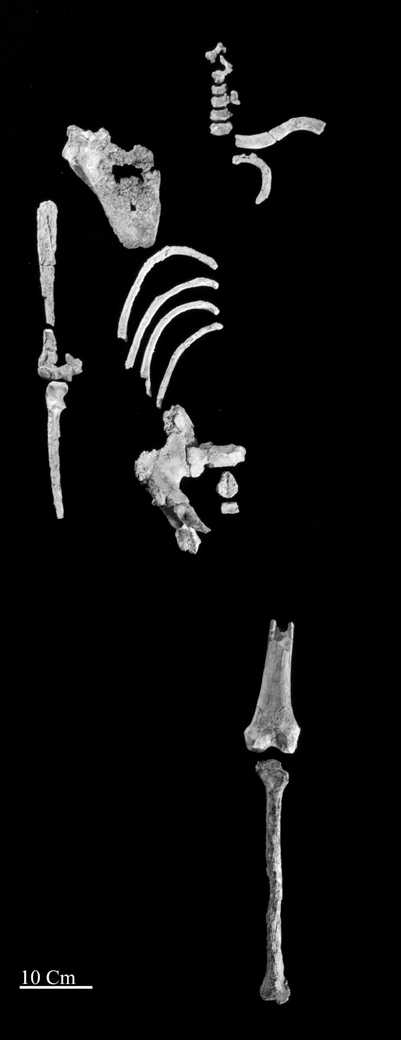
The long awaited publication of the partial hominin skeleton from Woranso-Mille, Ethiopia (shown on the left) should send shock waves through paleoanthropology. Yohannes Haile-Selassie et al. describe the specimen, which is dated by radiometric and paleomag techniques to 3.58 mya, in the on-line edition of PNAS now available. Older than “Lucy” by nearly 400 ky and slightly younger than the Laetoli footprints, the specimen includes key skeletal elements that clarify the anatomical adaptations for bipedality in early australopith hominins. According to the authors they are surprisingly modern, supporting contributing author Owen Lovejoy’s long-standing argument that A. afarensis had a modern striding gait and had lost functional adaptations for arboreality. At nearly 3.6 mya the Woranso-Mille hominin is less than one million years removed from Ardipithecus ramidus (~4.4 mya), but shows a near modern upper and lower limb morphology. Ar. ramidus on the contrary shows an extremely primitive post-cranial anatomy that is in many ways more Proconsul-like than modern ape-like, with well-developed arboreal adaptations. It would require a very quick evolutionary makeover to account for the necessary transformation of Ar. ramidus into Au. afarensis as described in the paper. Before this publication the degree of bipedalism that the partial skeleton illustrates was thought to be restricted to the genus Homo. If we reject the ancestor/descendant relationship between Ar. ramidus and Au. afarensis, where does that leave Ardipithecus as a hominin? It is most likely that Ar. ramidus represents a relic species, phenetically little removed from the last common ancestor of the African apes and humans. This conclusion thus leaves unanswered the question, from whence came Australopithecus?
International Symposium on Paleoanthropology in Commemoration of the 20th Anniversary of the Discovery of the Skulls of Yunxian Man
I attended an international conference celebrating the 20th anniversary of the discovery of “Yunxian Man” held in China from June 14-17. “Yunxian Man” is known from two complete crania (skulls without the lower jaw) found and excavated between June 1989 and June 1990 in the central Chinese province of Hubei. It was announced at the conference, held in Yunxian City, that the skulls have been re-dated to nearly 1.0 million years ago by Chinese scientists.
At the Conference I compared the new age attributed to “Yunxian Man” with the 1.0 million year old date given to a previously unknown human mitochondrial DNA (MtDNA) lineage announced earlier this year by German scientists at the Max Planck Institute for Evolutionary Anthropology in Leipzig. The team, led by Johannes Krause and Svante Pääbo, sequenced ancient mitochondrial DNA from a finger bone of a hominin found in Denisova Cave in southern Siberia. While the specimen comes from a population that lived about 30,000 to 48,000 years ago in the Altai Mountains in Central Asia a detailed analysis of the Denisova mitochondrial genome shows that it shared a common ancestor with modern humans and Neanderthals about 1.0 million years ago. Other recent research at the same Institute provides evidence that modern and archaic humans (including Neanderthals) had interbred on first contact.
Prof. Li Tianyuan, the principal investigator of “Yunxian Man,” and I gave papers at the Chinese symposium detailing how the Yunxian skulls exhibit a mix of features seen in both Homo erectus and H. heidelbergenis. H. heidelbergenis is thought by many scientists to be the common ancestor of both Neanderthal Man and modern humans. I proposed that given its age and physical features “Yunxian Man” is an ideal candidate for the 1.0 million year old common ancestor of H. heidelbergenis and the Denisova hominin. This would place the origins of H. heidelbergenis in Asia rather than Africa as previously thought.
Dr. Wu Xinzhi, the dean of Chinese paleoanthropologists, who chaired the meeting, gave a paper in which he presented evidence of Neanderthal-like traits in later human fossils from China, suggesting that Neanderthals, known to have lived near Denisova Cave, spread south into China interbreeding with indigenous Chinese hominins. This supports the idea that human evolution has been characterized by a complex web of interactions between multiple incipient human lineages that hybridized with one another on first contact, spurring the evolutionary events that led to the emergence of modern H. sapiens and other hominin species.
In keeping with these new developments I proposed a “Multiple Dispersal” model for human evolution more in keeping with recent genetic evidence than the earlier Multi-regional or Out of Africa Replacement models. In the Multiple Dispersal model humans evolved into distinct species after leaving Africa nearly 2.0 million years ago. Some of these species subsequently spread into adjacent regions hybridizing with the hominins they encountered. The westward dispersal of Yunxian Man’s descendents eventually gave rise to archaic humans in Europe (H. heidelergensis) and Africa (H. rhodesiensis), which then evolved respectively into the Eurasian Neanderthals (H. neanderthalensis) and the African ancestors of modern H. sapiens. New genetic evidence suggests that modern humans then spread out of Africa 60,000 years ago meeting up and hybridizing with Neanderthals in the Near East and the archaic descendents of H. yunxianensis in East Asia.
Other papers presented at the conference discussed evidence for an early appearance of modern H. sapiens in China. Dr. Shen Guanjun, who has re-dated many Chinese hominin sites using advanced radiometric techniques, said that a number of Chinese sites yielding modern human teeth were older than 100,000 years in age, tens of thousands of years before they are supposed to be there according to the Out of Africa theory of modern human origins. In a following paper Dr. Liu Wu showed how these teeth had typical East Asian features still seen in today’s Asian populations. I proposed that this evidence supported the likelihood of an early dispersal of modern humans out of Africa which hybridized with local archaic Chinese. A second wave of moderns coming out of Africa reached China around 45,000 years ago They then mixed with the earlier hybridized Asians giving rise to the Asian populations of today. Similar patterns most likely also occurred in Europe and elsewhere, lending further support to a Multiple Dispersal model for human evolution.
Friday, June 11, 2010
A Multiple Dispersal Hypothesis for Interpreting the Pattern of Hominan Evolution in China
The Yunxian hominans are represented by two complete crania discovered in 1989 in Yun County (Yunxian), Hubei Province. Although crushed and distorted to varying degrees they are the most complete early middle Pleistocene hominan crania known. They preserve intact the face, cranial vault and cranial base and as such are extremely important materials for assessing the amount of variation in local paleodemes, and regional populations of Pleistocene hominans. They also serve as an invaluable resource for evaluating the amount and character of polytypic variation in archaic hominans. A conference celebrating the 20th anniversary of the discovery of the Yunxian crania will be held in China in early June. I'll be presenting a paper at the conference proposing a multiple dispersal hypothesis for interpreting hominan evolution in China and elsewhere.
I'm baaaaaaaaaaack!!!!!!!!!!!!!!!!!
For all you loyal followers (1), I'm back. I can't rightly recall why I suspended operations in 2006. Probably just felt it was too big a burden with too little a return. But I've since realized that I actually may have a slight presence on the web so maybe the effort is worth while after all. Since then I've had a few narrow escapes with the grim reaper, heart surgery, an automobile accident, kidney failure and a ruptured gall bladder, but I ingratiated myself to the hospital staff and they gave me a reprieve. Better to thank your doctors than thank "god". Anyway, now that I'm back on track and about to leave for China for an "International Conference on The Yunxian Hominids," celebrating the 20th anniversary of their discovery in 1989, I feel that it's an appropriate time to rev things back up.
A lot has gone on in China over the last four years. I'll try to update this site so it can serve as a repository for posts dealing with new developments (and a review of older developments) in the paleoanthropology and paleontology of China and East Asia.
![]()
